|
|
|
Commentary on this paper will be posted at a later date. Estimating Ethnic Genetic Interests: Is It Adaptive to Resist Replacement Migration?Frank Salter (Max Planck Society, Human Ethology, Von-der-Tann-Strasse 3, D-82346 Andechs, Germany; e-mail: Salter [at] humanethologie.de) Population and Environment, Vol. 24, No. 2, November 2002. Abstract: Analyses of the costs and benefits of immigration have not considered the dependence of an ethny’s reproductive fitness on its monopoly of a demarcated territory. Global assays of human genetic variation allow estimation of the genetic losses incurred by a member of a population when random fellow ethnics are replaced by immigrants from different ethnies. This potential loss defines an individual’s ethnic genetic interest as a quantity that varies with the genetic distance of potential immigrants. W. D. Hamilton showed that self-sacrificial altruism is adaptive when it preserves the genetic interests of a population of genetically similar individuals. Ethnic genetic interest can be so large that altruism on behalf of one’s ethny—‘ethnic nepotism’—can be adaptive when it prevents replacement. It follows that ethnies usually have an interest in securing and maintaining a monopoly over a demarcated territory, an idea consonant with the universal nationalism of Bismarck and Woodrow Wilson. Key Words: genetic interests; replacement migration; immigration; ethnicity; Hamilton’s Rule; inclusive fitness; evolution; territory; carrying capacity; kinship. INTRODUCTIONDoes ethnic competition over territory pay off in terms of reproductive fitness? The question is barely raised in contemporary analyses of population, even when discussing the costs and benefits of immigration on such a scale that it is appropriately called ‘replacement migration.’1 Perhaps the idea that humans have reproductive interests does not occur to most social scientists after almost a century of that tradition separating itself from the biological sciences (Salter, 1996). Whatever the cause, analysts behave as if the only interests humans have are ‘proximate,’ such as economic and physical security, rather than ‘ultimate,’ in the form of genetic continuity. Thus it is implied that immigration on any scale is acceptable so long as it raises aggregate income or makes life more interesting (e.g., Rubin, 2000; Simon, 1989). The focus on proximate interests raises doubts from the perspective of ethnic nepotism theory (Eibl-Eibesfeldt, 1972; van den Berghe, 1981). According to this theory, ethnic solidarity is due to an ethnic group—hereafter referred to as an ethny2—constituting an extended family. From this perspective accepting replacement migration for the convenience afforded by migrant labor is equivalent to foregoing bearing children in order to live in greater luxury. But this argument has not as yet been supported by quantification of ethnic genetic interests—the number of copies of an individual’s distinctive genes carried in his or her ethny. Without such estimation it is impossible to determine who has it right, those who see ethnies as extended families, or those such as Lewontin (1972) who deny that humans have genetic interests in their ethnies. In this paper I shall not question the verities of classical liberalism—that international trade can produce win-win outcomes and that wars should be fought only in defense and as a last resort when diplomacy fails (Caton, 1988). But I shall disagree with the view that the benefits of interethnic trade necessarily render mass immigration harmless to native ethnic genetic interests. I begin with a qualitative argument based on evolutionary theory before introducing quantitative theory and data from population genetics. From the neoDarwinian perspective the issue is whether being altruistic towards one’s ethny can be adaptive in the context of ethnic rivalry. Behavior is adaptive when it maintains or increases the frequency of one’s distinctive genes in the population. Rivalry can involve peaceful as well as violent means. The latter consist of armed conflict, including warfare both defensive and aggressive. Examples of peaceful means are competitive breeding (Parsons, 1998; 2000) and discrimination, for example in controlling borders and in economic affairs (Bonacich, 1973; Landa, 1994; Light & Karageorgis, 1994). In neoDarwinian theory, genes are the basic unit of selection and humans are ‘survival machines’ evolved to perpetuate them into succeeding generations (Dawkins, 1976). Conversely, for humans and all other organisms, reproductive interest consists of perpetuating their distinctive genes (Hamilton, 1964). ‘[H]umans like other organism[s] are so evolved that their “interests” are reproductive. Said differently, the interests of an individual human (i.e., the directions of its striving) are expected to be toward ensuring the indefinite survival of its genes and their copies, whether these are resident in the individual, its descendants, or its collateral relatives. . . . ’ (Alexander, 1995/1985, p. 182). Thus genetic interests are the number of copies of our distinctive genes carried by reproducing individuals. Individual genetic interest is the number of copies carried by offspring. Familial genetic interest is carried by close kin, and ethnic genetic interest by one’s ethnic group. Genetic interests are often confused with ‘inclusive fitness.’ The latter concept was coined by Hamilton (1964) to describe his theory of altruism. It refers to the effect that an individual has on the reproduction of his distinctive genes, not to a static gene count. Thus a parent who has ceased reproducing and aiding her many offspring has no individual fitness. The two concepts are closely related, since an individual with positive fitness is acting so as to perpetuate his distinctive genes. Which modern events affect ethnic genetic interests? This interest is ultimately a matter of population size, which can be directly reduced through warfare, genocide, and the loss of limiting resources such as territory. The fact that a 30 percent loss of population is a 30 percent loss of ethnic genetic interest is obvious. But competition can have powerful effects without any behavior that is aggressive in the usual sense of the word. The prime example in the contemporary world is peaceful migration between states and high rates of reproduction by one ethnic group within multi-ethnic states. Like the bands and tribes in which humans evolved, states are territorially based and act to police their borders. The special quality of a defended territory is that it insulates a population from the vicissitudes of demographic disturbances in the metapopulation, namely the connected phenomena of uneven population growth and migration. When an ethny controls the borders of a territory that is large enough to support the population, loss of fitness relative to other ethnies is not necessarily fatal; it need not lead to replacement. A decimated, defeated, or impoverished population can quickly recover if it retains control of its territory, but a large-scale influx of genetically distant immigrants has the potential permanently to reduce the genetic interests of the original population. Territory adequately defended guarantees continuity and the chance to ride out a temporary downturn in numbers relative to other populations. The territorial component of the tribal strategy was so fundamental in Homo sapiens’ evolutionary past that it has become deeply imbedded as a psychological need. It has been long known from cross-cultural comparisons that all hunter-gatherer societies defend their territories against incursions by neighbors (Service, 1962; van der Dennen, 1995, pp. 427–8; 564–5). An innate component is indicated by the territoriality of humanity’s closest relatives, chimpanzees, who patrol the community borders (van der Dennen, 1995, p. 159). Among humans, territorial bonds and the sense of collective ownership vary in intensity and according to culture, for example both being strengthened by tribal rituals that cause individuals to identify and bond with the group’s territory (Eibl-Eibesfeldt, 1989, pp. 321–34). Whether the association of peoplehood with a land is genetically or culturally transmitted, it appears to be universal in both the tribal and national worlds (Connor, 1985; Spicer, 1971; see review by van der Dennen, 1995, pp. 427–8, 564–5). Persistent ethnic identities all include a territory as part of their identities, either presently or once occupied by the group (Spicer, 1971, p. 798). The evolutionary background of human tribal territoriality was discussed by Keith (1968/1947, chapter 4). In tribal societies a territory is necessary for subsistence and also for continued solidarity. Removed from its land, a tribe’s social organization tends to breaks down, reducing its ability to maintain independence and continued existence (p. 33). Keith argued for an intimate connection between a people as a descent group and their claim to a territory as essential constituent elements of nationhood:
Keith (1968/1947, p. 317) also recognized the psychological continuity between tribalism and nationalism. “Group spirit, tribal spirit or tribalism, national spirit or nationalism are one and the same thing. . . .” Loss of fitness within one’s own territory robs the native ethny of the time needed to recover numbers, mobilization, or organization, all contributing to a loosening of the ties between political leadership and ethny. Since in modern societies the state has come to replace traditional tribal institutions, loss of state sponsorship is likely further to undermine mobilization and organization. In a competitive world an ethny’s loss of ability to mobilize and organize as a self-interested group is tantamount to loss of fitness. Mass migration between diverse populations combined with the existence of collective goods in wealthy societies such as low cost medical support and other forms of welfare have produced effective ethnic competition within many Western states. For example the founding European-derived ethnies of the United States, Canada, Australia, Britain and some other Western societies are declining as proportions of the overall population due to periods of liberal immigration policy from the 1960s. These challenges are real enough for majorities, but minorities have usually fared worse, as diaspora peoples have discovered through the centuries. Not to control a territory creates risks of repeated group subjugation, displacement, and marginalization. For all of past human experience and still today, territory is a resource for maintaining ethnic genetic interests in the long run. The foregoing qualitative argument might be plausible, but only a quantitative analysis will allow us to assess whether, and under which circumstances, immigration harms native populations’ genetic interests to a significant degree. Such an analysis requires clarification of territorial carrying capacity and the quantification of ethnic kinship. CARRYING CAPACITYThe vulnerability of ethnic genetic interests to immigration depends on there being some limit to population growth that causes immigrants to displace native born. Hardin (1968; 2001/1974; 1993) argues that in the modern world most habitable spaces have been colonized. Moreover, the earth’s surface has a carrying capacity, as do its parts. This is the maximum population beyond which some value, such as freedom from hunger or overcrowding, is lost. The most basic carrying capacity is the number beyond which population growth is self correcting, because any further growth is cancelled out by die-offs. Technological advance can increase carrying capacity, but not indefinitely. E. O. Wilson (2002) uses a different analysis to make essentially the same point: the earth is full and its present population is probably unsustainable. His formulation is based on the ‘ecological footprint,’ ‘the average amount of productive land and shallow sea appropriated by each person in bits and pieces from around the world for food, water, housing, energy, transportation, commerce, and waste absorption’ (p. 72). The developing world has a per capita ecological footprint of about one hectare, while that for the United States is 9.6 hectares. If every human being were to consume at the average level of the United States with existing technology, four more planet earths would be needed to accommodate them. Populations and levels of consumption appear set to continue growing for the time being, but only at the cost of lost biodiversity and a collapse in the earth’s capacity to renew ecosystems. Population must level off at some point, whether through design or accident. Either way, average family size must fall to that of zero population growth, about 2.1 children. Hardin (1974) points out that to control run-away population growth in the world requires that every state limit its population. However, a society practicing such self discipline is vulnerable to immigration, which tends to reduce the size of the native population. Hardin did not discuss ethnic genetic interests, but did remark that in this situation the cost of immigration would fall “most heavily on potential parents, some of whom would have to postpone or forgo having their (next) child because of the influx of immigrants.” Hardin thus only considers individual fitness. Yet, as we shall see, the damage immigration inflicts on genetic interests contained in the native ethny is usually much greater than that contained in any family. Immigrants are liable to affect a country’s capacity to hold the native population. If the immigrants contribute to the economy in ways that the native population cannot, the carrying capacity is raised. If they are a drain on resources or even of average productivity, they lower that capacity by taking the place of potential native born. In the present example, let us assume that immigrants have equal capacities to the native born, and let us consider immigrants in lots of 10,000. This number of immigrants will lower the effective carrying capacity of a country by 10,000, more or less—more if the immigrants have a higher birth rate than the native population and less if their birthrate is lower. To simplify further, assume that birthrates are equal, in which case the loss of effective carrying capacity is 10,000. If the immigrants and native born have the same ethnicity the native population loses no ethnic genetic interests. Ethnic kin are being replaced by ethnic kin of similar degree. But if the immigrants are from different ethnic groups, especially genetically distant ones, there will be a loss of genetic interest for each member of the native population. How large is that loss? Answering this question will allow an assessment of immigration’s impact on ethnic genetic interests and, potentially in turn, its impact on society and politics. GENETIC INTERESTS AND KINSHIPThe scale of ethnic genetic interests can in principle be calculated from the data of population genetics, the science of counting and explaining gene frequencies. The quantity of direct relevance is the coefficient of kinship. This was commonly used until the 1980s, and is of special utility in genetic epidemiology (e.g., Morton, 1982; Gudmundsson et al., 2000). However, kinship coefficients are limited to within-population measures, or within a regional cluster of populations. What is needed for present purposes is a global assay of inter-population kinship measures, to capture the effect of immigration from adjacent and distant ethnies. Some ethnic groups are closely related, originating from the same regional or continental population. Other groups are more distantly related. Unfortunately, no world assay of genetic kinship at the population level exists. A global genetic assay performed by Cavalli-Sforza et al. (1994) measured not kinship but variance or ‘genetic distance’ (Cavalli-Sforza et al., 1994, p. 29). Fortunately, there is a well defined relationship between genetic kinship and variance that can answer our question. In population genetics the coefficient of kinship, f, between two individuals is defined as the probability that an allele taken randomly from one will be identical to an allele taken at the same locus from another. This definition is close to that of Hamilton’s (1964) original coefficient of relatedness r, which he used in his classic formulation of inclusive-fitness theory (except that it omits reference to the alleles being “identical by descent” and in simple cases 2f = r). Modern population genetics theory usually refers to kinship rather than relatedness, since the latter is not very clearly defined mathematically. Hamilton subsequently redefined inclusive fitness such that genealogical relatedness—the basis of kin selection—becomes a special case of his theory. The new definition allowed kinship to be calculated from assay data alone, with no requirement for data on genealogical relationships. Genetic distance, FST, is, then, both a measure of genetic variance and a measure of kinship (Cavalli-Sforza & Bodmer, 1999/1971, pp. 399, 451; Cavalli-Sforza et al., 1994, p. 29; Wright, 1951). On this basis, Harpending (1979, p. 624) found that kinship can be expressed in terms of variance thus: fo = FST+ (1 - FST)[-1/(2N - 1)] where fo is the local kinship coefficient, FSTthe variance of the metapopulation, and N the overall population. When N is large, as it usually is with modern ethnies, a good approximation for the above equation becomes, simply: fo = FST. This formula allows the estimation of average kinship within local populations based on FSTmeasures. The kinship of two randomly chosen individuals in a population is zero. In the same context, two siblings have a kinship of 0.25. Hamilton’s original genealogy-based coefficient of relatedness measures siblings as sharing 0.5 of their genes, but Hamilton later (1970; 1972) redefined relatedness as a statistical measure of genetic similarity (Pepper, 2000, p. 356). Using this new definition individuals can have negative kinship, meaning that they share fewer genes than is typical for the population, as well as positive kinship, when they share more genes than is typical. (Therefore it is necessary to distinguish kinship from relatedness-by-descent.) Hamilton summarized thus:
Moreover, Michod and Hamilton (2001/1980, see especially pp. 108–9) reviewed alternate definitions of kinship formulated to that date and concluded that they were all equivalent. Thus inclusive fitness theory is not limited to explaining altruism between genealogical relatives, but between any individuals who have positive kinship as defined above, even if their relatedness is remote and cannot be documented. This frees the analyst from the “identical by descent” clause, allowing the direct analysis of inclusive fitness using genetic assay data alone. Without this broader conception of inclusive fitness direct analysis is impossible because genealogical data are very limited. Harpending explains the implication of his conversion equation thus:
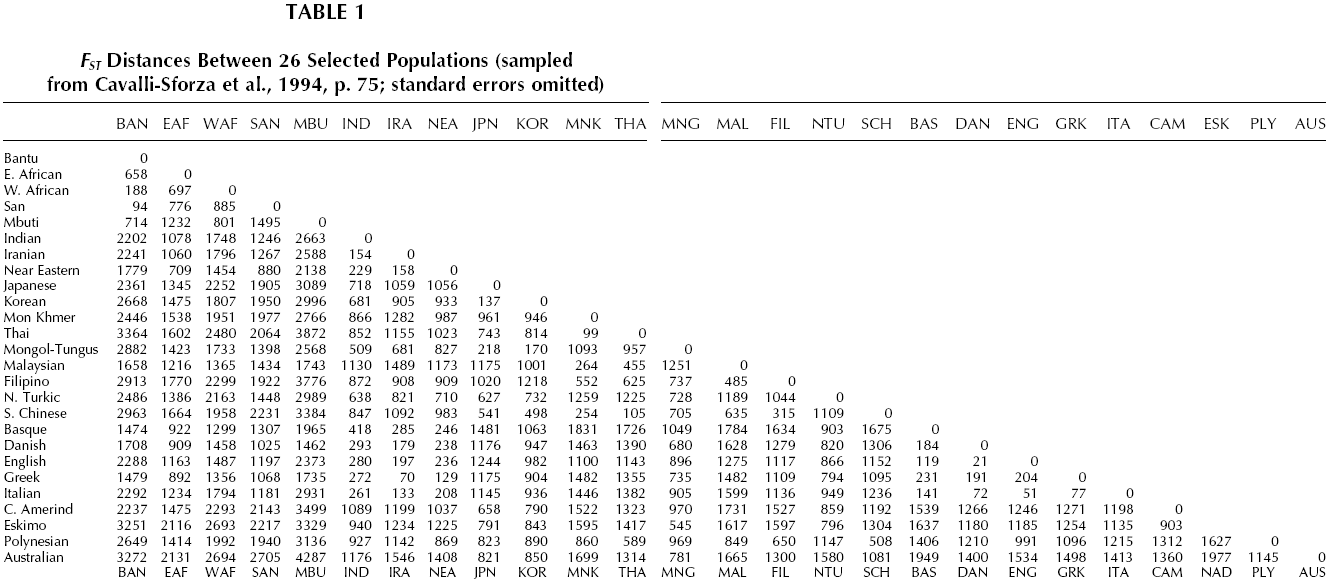
Returning to Harpending’s formula, this can be summarized by the rule that genetic variation between populations is equal to kinship within them. Kinship within an ethny thus varies in proportion to the genetic variation between it and competing ethnies. The resulting relativity of ethnic kinship can be illustrated with a version of Haldane’s (1955) famous hypothetical example of altruism between kin. Recall that Haldane suggested that it is adaptive to give one’s life to save two drowning siblings or eight drowning cousins. What he omitted to make explicit was the background assumption that the relatedness between random pairs in the population is zero. Cousins have a relatedness of one eighth not in an absolute sense, but in comparison to this zero background relatedness. Hamilton defined the kinship of two random members of a population as zero, but as we have seen when the variance of the meta-population is taken into account it is possible for relatedness of random pairs within a population to be more than zero. Hamilton (1975) argued that intra-group relatedness could rise as high as 0.5 (equivalent to a kinship coefficient of 0.25), which is the level of siblings. Families within such a population would then have even higher relatedness. What if the world consisted of cousins? For the purpose of estimating genetic interests, their kinship would be zero, and it would not be adaptive for them to show altruism towards each other. In this hypothetical case, there is no competing group against which cousins have an elevated level of kinship. Adaptive altruism would then be limited to the nuclear family, where relatedness is four times higher than between cousins (eight times higher for identical twins). If the world consisted of only one ethny, the relatedness of random pairs would be effectively zero for the purposes of estimating genetic interests; only genealogical kin would have higher kinship. Thus adaptive altruism would only exist between parents and offspring, siblings, cousins, etc. But in fact the world consists of a great many ethnic groups. Taking the whole world population together, the kinship of random pairs is zero. The question is, in this situation what is the kinship of random pairs chosen from an ethnic group? To answer this question we must turn to the data on genetic variance between populations. IMMIGRATION, GENETIC DISTANCE, AND LOSS OF GENETIC INTERESTData from Cavalli-Sforza et al. (1994, p. 75; see Table 1) indicate that intra-ethnic kinship coefficients range from 0.0021 (English in relation to Danes) to 0.43 (Australian Aborigines in relation to Mbuti Pigmies of Africa). In general, kinship between autochthonous populations within the one region is close while between continents it is distant. Using the two extreme cases just mentioned, Table 1 shows kinship in the following way. Recall from the discussion in the previous section that if the world population were wholly English then the kinship between random pairs would be zero. But if the world consisted of the English and Danes, then two random Englishmen would have a slightly positive kinship of 0.0021 (kinship coefficients are multiplied by 10,000 in Table 1). This is slightly closer than the kinship of eight linear generations separation, or a descendant to his or her great great great great great great grandparent. This is another way of saying that the English and Danes are very close kin. Australian Aborigines and the Mbuti people of Africa are very distant kin according to Cavalli-Sforza’s data. In a population made up of Mbuti and Aborigines, two random Aborigines are almost as closely related as identical twins, with a kinship of 0.43 (identical twins have a kinship of 0.5).
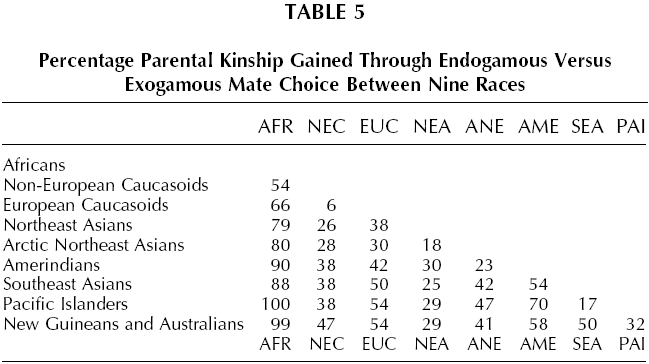
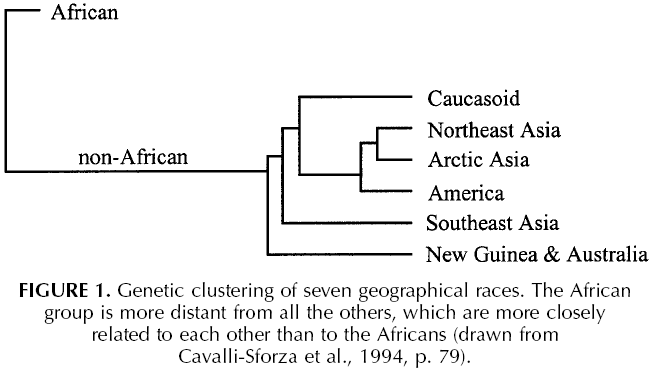
Taken together with the limits imposed by carrying capacity, these kinship coefficients mean that, other factors being equal, immigration is more harmful to the receiving population’s genetic interests the more genetically distant the immigrants. The FST distances shown in Table 1 allows us to estimate the loss in genetic interest caused by uni-directional migration of 10,000 individuals between 26 native populations. Appreciation of the genetic interests involved is aided by converting this loss to child-equivalents. Losses of genetic interest will not be counted in units of random fellow ethnics, but in the larger unit of offspring. In Hamilton’s (1975) town model the kinship of random co-ethnics could rise as high as that between parent and child in outbred populations. Hamilton noted that when this happened, actual parent-child kinship would rise significantly higher, though he did not specify how high. Harpending (2002) offers a formula for that higher figure, based on the genetic distance (FST) between populations. f = 0.25 + 3FST/4 Applying this formula allows us to estimate the impact on the genetic interests of a random Englishman of 10,000 ethnic Danes replacing 10,000 ethnic English (or vice versa). To simplify, let us assume that this is a neat replacement, so that over succeeding generations all the immigrants survive to reproduce.3 We also assume that the Englishman loses no genealogical kin in the process. Replacement involves two effects, the removal of 10,000 Englishmen and the introduction of 10,000 Danes, who in the mathematics of population genetics have negative kinship to the English population. Removal of the English in this case reduces genetic interests by 10,000 X 0.0021 = 21. The replacing Danes bring a negative kinship of the same magnitude. Subtracting the latter from the former gives a loss to a random Englishman’s genetic interests of 42 units. Now we express those units in number of children by dividing by the parent-child kinship of the English, which is f = 0.25 + (0.0021 x 3/4) = 0.2516 The number of children lost due to the immigration of 10,000 Danes is therefore 42/0.2516 ~ 167 children (or siblings). This is a large family indeed. Repeating the scenario with Bantu immigrants, the loss to a random Englishman’s genetic interests of replacement of 10,000 English is 10,854 children (or siblings).4 Bantu suffer the same loss from 10,000 English immigrants to a Bantu territory. This last figure is puzzlingly high. How can the loss due to replacement exceed the number replaced? Actually it does not, because we are counting genes, not individuals. The result only looks strange when gene counts are converted into child-equivalents. Random members of an ethnic group are concentrated stores of each other’s distinctive genes, just as children and cousins are concentrated stores. Some ethnies are so different genetically that they amount to large negative stores of those distinctive genes. Also, migration has a double impact on fitness, first by reducing the potential ceiling of the native population, and secondly by replacing those lost individuals’ familiar genes with exotic varieties. Referring to carrying capacity also helps understand the dramatic loss of fitness caused by interracial immigration. In a large nation the loss of 10,000 fellow ethnics, say due to a natural catastrophe or war, could be made up in a generation, as the population rose to the country’s carrying capacity. Filling their places with immigrants reduces that loss by an amount equal to the immigrants’ kinship with the natives. When Danes replace English, the loss is almost completely reduced because the two ethnies are closely related. But when Bantu replace English (or vice versa) the loss is exacerbated, because these populations have negative kinship with one another. The loss is not diminished by somehow being spread across the entire ethny. For the native it is a collective loss in the same way that collective goods are shared without being diminished. It applies to every randomly chosen member of the native ethny, wherever he or she may live. Children per se are not lost. It is assumed that there will be the same number of children in the society. Neither is it a symbolic loss for people everywhere, like some humanitarian disaster. After all, the immigrants produce replacement children. For them the process of ethnic replacement increases fitness. The loss is limited to the native ethny in a very personal way. For a native woman it is equivalent to the loss of her children and grandchildren, for a native man it is equivalent to the loss of his children and grandchildren, though on a much larger scale. The magnitude of these ethnic genetic interests means that the loss is only slightly mitigated if these individuals’ own children are not replaced. It becomes clear from these data that ethnic genetic interests are usually very large compared to familial genetic interests. The mathematics on which these estimates rely would need to be in gross error for this not to be true. Indeed, for inter-racial immigration the losses would still be large if ethnic kinship were one hundredth the values estimated above. Neither would inaccuracies in other assumptions of the analysis necessarily alter the result by much. For example, the genetic loss occasioned by cross-racial immigration can be mitigated by the immigrants increasing the country’s long-term carrying capacity. I have assumed nil economic benefit (and cost), but the genetic loss will still be large even if the economic benefit mitigates half or even 90 percent of the loss. The result is also robust for all except radical modifications of the assumption of a neat replacement of natives by immigrants, since partial replacement still causes large losses of ethnic kin. Ethnies are indeed super families as van den Berghe argued. Although being more dilute stores of genetic interest than families, ethnies can number in the millions and so are often orders of magnitude more precious. If immigrants replaced one quarter of the English nation of approximately 50 million people, the remainder would suffer a very large loss even if their own relatives were not affected. If 12.5 million Danes and similar peoples moved to England, the genetic loss to the remaining English would be the equivalent of 209,000 children. The corresponding loss due to the same number of immigrants from India would be 2.6 million children, and due to Bantus over 13 million children. All these losses apply in the reverse direction, if there was mass English immigration to Northern Europe, India, or Bantu Africa. Large ethnic genetic interests make public charity and self-sacrificial heroism directed towards one’s ethnic group potentially adaptive. As we see from the above estimates, ethnic altruism is most adaptive when it aids fellow ethnics in the face of competition from genetically distant ethnies, such as those belonging to different geographic races. Figure 1 shows the relative genetic distances of the major races, based on a world-wide assay of 88 genes by Cavalli-Sforza et al. (1994). The FSTmeasures between nine races are shown in Table 2. Subjects were individuals who could trace their ancestry from indigenous populations, the autochthonous peoples that inhabited a region before the great migrations of the modern era began to mix geographical races from about 1600.
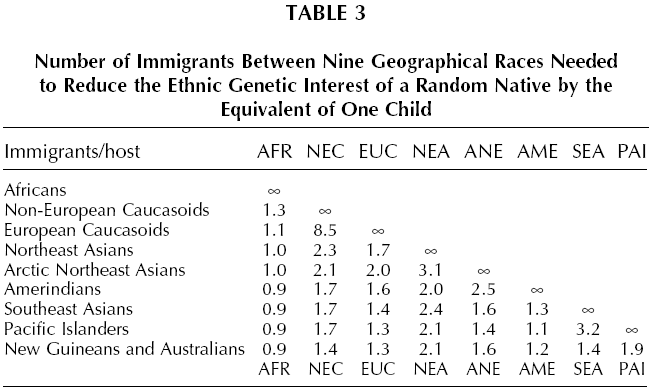
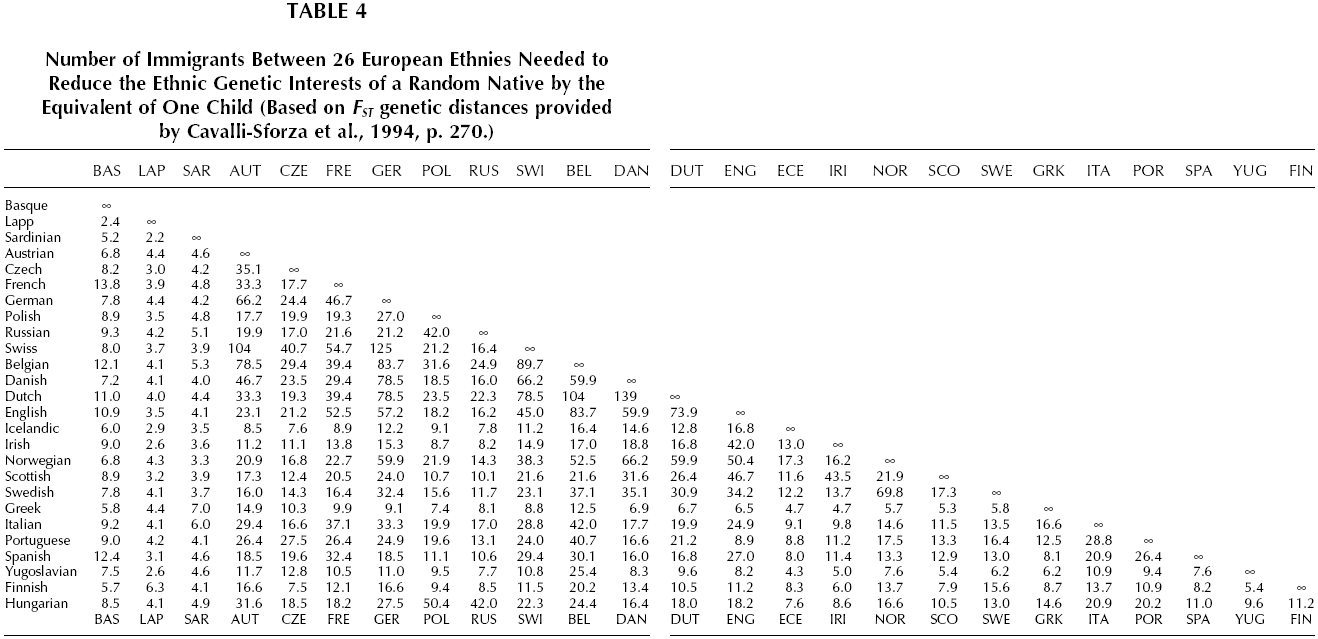
The genetic distance between English and Bantu is so great that, on the face of it, competition between them would make within-group altruism among random English (or among random Bantu) almost as adaptive as parent-child altruism, if the altruism were in the service of that competition. Thus it would appear to be more adaptive for an Englishman to risk life or property resisting the immigration of two Bantu immigrants to England than his taking the same risk to rescue one of his own children from drowning, unless the immigrants were bringing qualities of such economic value that they would permanently raise the Island’s carrying capacity. The same applies in the reverse direction, two Englishmen migrating to Bantu Africa constitute a greater loss of long-term genetic interest than does a random Bantu losing a child. But the genetic distance between English and Danes is so small that in the equivalent situation it would be adaptive to expend only slight altruism in resisting small numbers of immigrants. Despite the potentially large payoff, intra-ethnic altruism can be maladaptive when it benefits the reproduction of free riders within the group. This problem and institutional counter-strategies are discussed in Chapter 6. That cross racial immigration is most damaging to ethnic genetic interests becomes apparent if one considers the number of immigrants needed to reduce the ethnic genetic interest of a random native by the equivalent of one child (assuming as before that immigrants are economically equivalent to natives and that there is a finite carrying capacity for each country). Table 3 shows these numbers for immigration between races, Table 4 the numbers for immigration within Europe, a generally racially homogeneous region. The latter are usually about an order of magnitude greater than the former, meaning that immigration within a race is usually less harmful to ethnic genetic interests than immigration between races. There are exceptions. According to these data, immigration of non-European Caucasoids to Europe (and the reverse; see Table 3) causes less kinship loss than immigration of Lapps or Sardinians to England (or the reverse; see Table 4). Note, however, that this exception occurs within regional subsets of the broad Caucasoid racial group. The overall pattern is clear. Within local regions of Europe the genetic impact of immigration declines by up to another order of magnitude. Immigration between ethnies of the same race can still be maladaptive for the receiving population, but the threshold is typically 10 to 100 times that of inter-racial immigration.
Hamilton’s theory of inclusive fitness allows the calculation of, in principle at least, the number of coethnics that must benefit if an altruistic act is to be adaptive. Hamilton formulated a rule for calculating when an act of altruism may be adaptive, or ‘evolutionarily stable’, such that the altruist’s genes are not reduced in frequency in the population. Hamilton formulated his rule in terms of relatedness r, for which 2f is substituted here to retain compatibility with the data of population genetics. Hamilton’s Rule states that altruism is only evolutionary stable when b/ c > 1/2f where f is the average coefficient of kinship between the altruist and the recipients of the altruism; where b is the sum of fitness benefits to all individuals affected by the altruistic behavior; and where c is the fitness cost to the altruist. Hamilton characterized his rule thus:
Hamilton’s Rule eases the condition for adaptive ethnic altruism in the case of repeated altruistic acts of small cost to the giver, the more so when the benefit to the receiver is a multiple of that cost. For example, small change given to street beggars can have much greater value to the receiver than cost to the giver. Also, someone with discretionary control over hiring or awarding contracts can dispense large benefits at little or no personal cost. Altruism can be highly adaptive when it benefits a large number of fellow ethnics all at once. An act of charity or heroism that prevented 10,000 Danes from replacing 10,000 English would be adaptive even if it cost the altruist his or her life and with it all prospects of raising a family (at least a family of less than 167 children), since this would save the equivalent of 167 children for the altruist. Preventing replacement by 10,000 Bantu would warrant a much larger sacrifice because the genetic benefit is about 65 times larger. Despite these large benefits, ethnic altruism that does not contribute to security from replacement migration is maladaptive. The considerable genetic variation within all populations leaves much room for individual competition. Altruism is only adaptive when it serves to protect shared (ethnic) genetic interests. Ethnically competitive behavior easily obeys Hamilton’s Rule when the cost of ethnic competition is low, for example when competition takes the form of politics rather than sacrificing one’s life defending borders. An instructive example is the United States, the world’s largest recipient of immigrants. Since the immigration reforms of 1965, Americans of European descent—“whites”—have fallen rapidly in relative numbers. In 1960, the white population was 88.6 percent (Statistical Abstract of the United States: 2001, Table 10; Hispanics were not yet counted separately). By the 2000 census the non-Hispanic white population was down to 69.6 percent of the population (U.S. Census Bureau website, May 2002, Table PCT002). While the white population kept growing, the higher birth rate of minorities and the large immigration influx of almost one million legal immigrants per year plus many illegals, caused its proportion of the population to decline by 21 percent within two generations. By 2050 the Census Bureau projects the non-Hispanic white proportion of the population to be 52.8 percent (Statistical Abstract of the United States: 2001, Table 15). The United States’founding population is heading towards minority status by 2060. Note that the statistics just cited are approximations. The white population is somewhat larger than stated, because a minority of Hispanics are of predominantly European descent. It is impossible to put a precise figure on the racial makeup of Hispanics, because the necessary information is not collected by the Census Bureau. Immigration is effectively replacing large numbers of ethnic kin of the native born, not only whites but all native born Americans who are ethnically distant from the major immigrant ethnies; that includes African Americans. These populations have increased absolutely, but in the long run, when America’s carrying capacity is reached, they stand to be a much smaller proportion of the population than they were in 1965. Interbreeding will not change the situation, because what counts is the proportions of a people’s distinctive genes in a territory, irrespective of how they are mixed. By the same token, the immigrants now diluting the native-born gene pool are benefiting their own genetic interests. Their home countries typically accept no immigrants and have much higher fertility rates or populations than do Western societies. So the process is a boon for the genetic interests of immigrant ethnies. If one construes ethnically-restrictive immigration policy as a form of inter-group competition, then the American majority would have been handsomely rewarded in terms of genetic continuity had it engaged in competitive behavior of this form by maintaining the quota system put in place in 1924. This system was aimed at maintaining the ethnic proportions that existed within the country in the late 1800s. Conversely, efforts by minorities—meaning lobbying efforts within the country combined with instances of illegal entry, work, and residence—to overturn that quota system and widen the 1965 open door constituted acts of competition with the ethnic majority. Those efforts, combined with other lobbies such as some employers seeking cheap labor, has decreased white genetic interests more than all American war losses combined. If genetic interests are so valuable, why do countries such as the United States accept their loss on such a large scale? The short answer is that, as a rule, they do not. The United States is one of a handful of societies, all Western and mainly English-speaking, that are currently managing the replacement of a large fraction of their founding populations as a proportion of their territories’ carrying capacity. The immigration policies of the overwhelming majority of societies are slight variations on a total ban. Any explanation of the acceptance of ethnic replacement must therefore deal with particularities. What is different about America? One candidate explanation that can be immediately rejected is that the American majority’s failure to defend itself against genetic replacement reveals a preference for the dissipation of genetic interests or ambivalence about its value. This is an unlikely proposition, since it assumes widespread knowledge of the analysis being put forward in this paper. It also assumes that the electorate is biologically literate and is kept informed of events impinging on their genetic interests. I noted above that the US Census Bureau does not keep track of relative ethnic fitness within the country. This is odd from the Darwinian perspective. It is understandable that in a democracy precise statistics are kept of murders and accidental deaths, which number in the thousands, because such events not only affect individual fitness but are barometers of public safety. Yet the annual displacement of hundreds of thousands of native-born Americans is haphazardly reported. The pre-Darwinian state of western political culture is surely a contributing factor to the loss of relative ethnic fitness. A more hopeful explanation is that, despite ignorance of the biological issues at stake, United States policy has somehow managed to shape immigration so that it is not maladaptive, despite coming mainly from populations genetically distant from the European gene pool. As noted earlier, economic effects can indirectly benefit the native population by increasing carrying capacity thus helping all resident groups increase their absolute fitness. Has post 1965 immigration to the United States boosted the genetic interests of the native born? This issue is not treated by the extensive economic and sociological literature on immigration, but it seems likely that some immigrants have enhanced the majority’s fitness through provision of comparatively low-priced expert services. Others have helped undercut uneconomic labor costs by providing a pliable low-cost pool of labor, and stimulated those industries responsible for providing education, housing and other infrastructure. These benefits are compatible with many immigrants being a net burden on the public purse and the jobs of the native born (Borjas, 1999; Smith & Edmonston, 1997). Another set of benefits flows from the cultural diversity introduced by immigrants, though this too incurs costs in social capital, as unspoken ethnic ties are replaced by communal conflict and alienation. The United States is no longer an ethnic nation but a diverse multicultural state. This makes it more difficult to extract much public altruism in the form of civic altruism and patriotic sacrifice (Alesina et al., 1999; Walzer, 1990/1992, p. 24; see literature review by Salter in press-a). Even if immigrants always boosted genetic interests, this argument can be taken to absurd lengths. For example, if an immigrant ethnic group is generally more productive than the natives, complete replacement would result in the carrying capacity being greatly increased, though the natives would hardly benefit. Is an economy meant to serve people or be an end unto itself? Clearly there will be an optimum level of immigration rather than a maximum. And that maximum will be less than indicated by economics alone if the natives wish to maintain control over their affairs. The state is a powerful instrument for maintaining group identity, mobilization and coordination. In a democracy, maintaining possession of the state apparatus requires maintaining demographic dominance as well as control of the economy, even if that lowers the rate of growth. Any argument based on economic carrying capacity must guard against being short-sighted, since limits to growth can take generations to take effect. Eibl-Eibesfeldt (1998) has argued that humans are congenitally obsessed with short-term gains in modern environments due to their evolutionary history. Imagine that in 1600, before they sold Manhattan, American Indians had been offered an informed choice between two futures in the year 2000. One future was the present United States with a level of economic development unattainable without the efforts of millions of settlers and immigrants from Europe. The other was one or more Indian nations in possession of the present area of the United States but with economies less developed than at present. Which would they have chosen if they had valued ethnic genetic interests? A temporary delay, even one of decades or centuries, in acquiring some skill or institution would seem a weak excuse for sacrificing the future of all succeeding generations. And in the modern era of automation a rush to import manual labor would seem especially short-sighted. Even if the economy and with it the population temporarily receded, this would be a small price to pay for retaining a continent for one’s descendants. THE MALADAPTIVENESS OF UNITARY MULTI-ETHNIC SOCIETIES AND A PROPOSAL FOR UNIVERSAL NATIONALISMPopulation-genetic data show that great genetic diversity still exists between ethnic groups. Modern ethnies are less homogeneous than their ancestral tribal forebears due to improvements in transport and the increasing size of administrative and economic units. But the same developments have widened the competitive realm from neighboring tribes to encompass all human populations. Populations that have not been in contact for many tens of millennia are now able to migrate to each other’s lands in large numbers. Enculturation of immigrants does not necessarily eliminate ethnic competition. The genetic distance between modern ethnies is often so great that it is visible in racial differences of skin color, physiognomy, and body proportions. These differences are reliable markers of significant ethnic kinship distance, as predicted by Hamilton (1975, p. 144): “[A]t about the point where the colony members are related to each other like outbred sibs it should become relatively easy for individuals to detect a fairly clear difference in appearance when comparing fellow colony members with outsiders. . . . [This] should make possible fairly accurate separation of ‘us’ and ‘them’ . . . ” This is not the “green beard” effect in which the same gene causes altruism and some visible characteristic, since Hamilton is referring to “several traits which are independently inherited”. He is thus referring to the same kind of broad-based genetic similarities that characterize genealogical kin. Citizens in multi-ethnic societies, especially multi-racial ones, are faced with an invidious choice. They can do what comes naturally and practice ethnic nepotism, which is adaptive by promoting relative fitness but, especially when exhibited by the majority, engenders social conflict and can make the economy less productive. Alternatively, citizens can adopt the discipline of non-discriminatory behavior which, when they are in the majority might raise the carrying capacity of society as a whole but sacrifices relative fitness. Eliminating majority discrimination only worsens the problem if minority discrimination is not also eliminated, since asymmetrical mobilization is likely to give minorities disproportionate influence, hastening the majority’s decline. For historical reasons this asymmetry is commonly accepted as multiculturalism that is “working.” Until about 1965 Western multi-ethnic societies gave the ethnic majority precedence, disadvantaging minorities. Majority free-riding on minority labor was commonplace, for example in the institution of slavery and the post-emancipation importation of low-cost labor. However, since the 1960s and the broad acceptance of civil rights claims a new modus operandi developed. Majority ethnic groups restrained their own discrimination towards minorities more than the reverse, a formula known as multiculturalism. This unilateral withdrawal from ethnic competition arguably benefited the economy as a whole and certainly benefited minorities. But this formula was risky for a majority when minorities were genetically distant, since it turned minorities into potential free-riders on majority altruism. In several Western societies, including the United States, Canada, and Australia, the new approach coincided with the lifting of controls on non-European immigration and consequent rapid changes to those countries’ ethnic makeup. From the majority perspective, it would seem that the only thing more maladaptive than multiculturalism that does not “work” is multiculturalism that does “work.” Minority free-riding occurs in a number of ways. When there is ethnic stratification, a characteristic of all multicultural societies, minority freeriding can occur at the bottom of the class structure in the form of socialwelfare and other benefits conferred by public goods. A redistributive system thus entails majority-ethnic taxpayers paying for their own loss of relative fitness by financing reproduction by families belonging to other ethnies. Ethnic majorities can also find themselves economically or culturally dominated by a highly competitive minority, when minority free-riding is liable to take top-down forms such as steering cultural, immigration, and foreign policies towards minority goals with collateral harm to majority interests. Multi-ethnic societies thus tend to be maladaptive for majorities under multicultural regimes and maladaptive, or at least inequitable, for minorities under traditional regimes. Confirmation of the large scale of ethnic genetic interests will warrant a reconsideration of the essential nature of national interests as well as the meaning of a just world system. The present analysis reaffirms the value of national sovereignty. From a Darwinian perspective, a sovereign polity is one that controls immigration to its territory, both in times of war and peace. A biologically just world order might be something like the universal nationalism advocated in the nineteenth century by Otto von Bismarck and in the early twentieth century by Woodrow Wilson. In a growingly integrated world, one people’s disaffection is liable to reduce everyone’s interests, so that all stand to benefit from a formula that acknowledges both the need for autonomy and the reality of interdependence by respecting the most basic interest, genetic continuity. The same formula would make stabilization of the world’s population evolutionarily sustainable by protecting the genetic interests of the most restrained populations. Universal nationalism means thinking again about ethnic states where the state unambiguously serves the ethnic interests of the majority. By ethnic state I mean something closer to the traditional German than the French model of the nation. The German model adopts ethnicity as shared descent as a criterion of citizenship and thus offers a constitutional barrier to replacement migration. In the second half of the twentieth century this barrier collapsed in wealthy states that adopted the French model. The main risk appears to be the substitution of a set of abstract concepts as the defining symbols of the nation in place of ethnicity. The ‘constitutional patriotism’ espoused by anti-nationalists such as J. Habermas (1998) is a formula for reconciling ethnic majorities to their own demise while serving the sectional interests of minorities and free-riding elites. Every state currently managing the replacement of its founding ethnic group (e.g., Australia, Britain, Canada, France, USA) has adopted constitutional patriotism of one form or another. This is usually linked to the doctrine of multiculturalism, which encourages minority ethnocentrism while directing majority patriotic feelings towards universalistic ideals. According to this formula a country would lose nothing if the founding ethnic group were peacefully replaced so long as some set of values—democracy, equality, non-discrimination, minority rights—were retained. The combination of constitutional patriotism and multiculturalism is, as one would expect, subversive of the ethnic interests of the majority. The revival of ethnic nationalism would run counter to current liberal democratic opinion. However, from the original liberal perspective such a substitution would be warranted since it would salvage majority interests from precipitous decline. Minority interests could be partly served in an evolutionarily stable manner through federalism, in an evolutionarily uncertain manner through assimilation, as well as in a preventative manner through restrictive immigration combined with a foreign policy that adopted an informed and even-handed version of nineteenth century nationalist doctrine. I do not suggest that any simple or uniformly beneficial solution exists. Consider the question of the optimal size for nation states. For the purpose of conserving genetic interests, small is indeed beautiful, since a population will usually have a higher concentration of distinctive genes the smaller it is. But for the purposes of economic and military viability, bigger has usually been better. The traditional nation states that had developed by the end of the nineteenth century are actually amalgamations of ethnies. Although too large to represent a pure ethnic interest, states had to be large in an anarchic international system if they were to be economically and militarily viable. The spread of free trade zones has allowed small states to thrive, but if the model is to be generalized powerful global institutions will be needed to enforce international law as an alternative to war. International governance carries its own risk, including the entrenchment of an exploitative global elite unmoderated by alternate centers of power, and the plundering of the most productive economies by the least. Another factor certain to confound simple solutions is the mass immigration that has occurred over the last few decades, which has increased the number and the zero-sum character of conflicting ethnic interests. But there is hope in humankind’s ingenuity. For example, Eibl-Eibesfeldt (1989) has discussed ethnic federalism constrained by reproductive interests, while recognizing that no all-purpose solution exists.
Nation states have the advantage of allowing relatively individualistic behavior to be adaptive by muting ethnic genetic interests as a factor in intra-state politics and economics. Perhaps this is why the industrial revolution was the product of nation states in the European tradition. In principle the ethnic state is conducive to liberalism in its political and economic forms. When competition is limited to individuals and families and does not involve ethnies, individuals can behave in ways that approximate the optimal (individual) utility maximization modeled by econometricians. But in multi-ethnic states individual economic rationality is maladaptive when it leads to loss of relative ethnic fitness and ethnic stratification, as is common in multicultural societies. From an evolutionary perspective a theoretical problem remains with ethnic states. A homogeneous nation state precludes ethnic free-riding on collective goods, but is no guarantee against free-riding by co-ethnics. Intraethnic elite free-riding should be less maladaptive than inter-ethnic freeriding, if only because co-ethnics benefit in the former case; but it still endangers the inclusive fitness of the majority of the ethny as well as the inclusive fitness of non-free-riders relative to co-ethnics. Thus altruism directed towards the ethny is evolutionarily unstable when free-riding is not controlled. The two main types of non-ethnic free-riding parallel the types of ethnic free-riding: welfare-dependent classes and elites. A democratic state will lose legitimacy if it fails to provide welfare by transferring resources from rich to poor. However, a modern economy will lose its competitive edge if it does not reward large economic contributions with substantial advantages in prestige and resources. A related phenomenon is the managerial class, critical for running the state and corporate bureaucracies. This elite group has privileged access to the levers of cultural and political power and tends to develop self-serving ideologies and institutions that put its interests ahead of the national interest (Burnham, 1975/1964; Francis, 1999/1984). Class and ethnic interests potentially conflict because they threaten defection by winners as well as losers in the competition for resources. Class competition is complicated by reproductive interests at the family level, in contrast to ethnic competition which involves group-level interests. The problem is to strike a biosocial contract between the classes that does not compromise individual reproductive opportunities while defending jointly held ethnic kinship. If a solution exists to this problem it will probably involve treating ethnic genetic interest as a collective good (Goetze, 1998), one that is managed by the state as an evolutionary group strategy (Boyd & Richerson, 1985; MacDonald, 2002/1994). Collective goods are already managed by states, such as group defense, education, and communication infrastructure. Contemporary states also exercise some control of free-riding on these collective goods through such means as law-based policing of public behavior, universal taxation, and compulsory national service. One challenge would be to maintain the transparency of both the strategy and its management by the state as a means of protecting against elite free-riding of the form that manipulates state power. An equally important challenge would be to prevent the double-edged sword of ethnic nepotism from cutting both ways. This will involve such strategies as balance-of-power diplomacy and participation in international institutions that blunt aggressive military nationalism in the international arena. The trick is to advance this agenda while leaving room for other institutions that husband national solidarity and its multiple benefits (Salter 2002; in press-b). This would seem the appropriate domain for constitutional idealism in which abstracted values become a basis for legitimating social arrangements. A principled resolution of class and national conflict could become the basis for a truly universal nationalism. ACKNOWLEDGMENTSI am indebted to Henry Harpending for coaching me in the application of his formula and to Jim Bowery, Rick Michod, and Vince Sarich for critiquing it. Alan Rogers provided the same formula in a similar mathematical proof. ENDNOTES
REFERENCES
| |

Existential IssuesWhite Genocide ProjectOf noteRecent CommentsAlso see trash folder. Graham_Lister commented in entry 'Using high school kids and their little "R word" friends for the Repressive Tolerance agenda' on 02/21/12, 03:00 PM. (go) (view) Leon Haller commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 02:29 PM. (go) (view) Lee John Barnes commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 12:42 PM. (go) (view) anon commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 11:59 AM. (go) (view) Gudmund commented in entry 'Population Matters' on 02/21/12, 11:56 AM. (go) (view) Graham_Lister commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 09:53 AM. (go) (view) Graham_Lister commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 09:14 AM. (go) (view) Lee John Barnes commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 08:04 AM. (go) (view) antizionist commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 07:51 AM. (go) (view) Sanatana Dharma commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 07:16 AM. (go) (view) james cruze commented in entry 'Giving up (our) liberty for (Islamic) security' on 02/21/12, 06:47 AM. (go) (view) Guessedworker commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 05:24 AM. (go) (view) Graham_Lister commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 05:15 AM. (go) (view) Bill commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 03:58 AM. (go) (view) Captainchaos commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 03:01 AM. (go) (view) Dan Dare commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 02:04 AM. (go) (view) Captainchaos commented in entry 'The Birth of the 'New-Man' under Liberalism' on 02/21/12, 01:53 AM. (go) (view) Captainchaos commented in entry 'The Birth of the 'New-Man' under Liberalism' on 02/21/12, 01:41 AM. (go) (view) Captainchaos commented in entry 'Population Matters' on 02/21/12, 01:06 AM. (go) (view) Captainchaos commented in entry 'Kai Murros: Moscow Speech 2010' on 02/21/12, 12:30 AM. (go) (view) Leon Haller commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 11:44 PM. (go) (view) dc commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 11:43 PM. (go) (view) dc commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 11:41 PM. (go) (view) Williams commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 11:22 PM. (go) (view) Gary commented in entry 'More on the Indian beauty question' on 02/20/12, 10:59 PM. (go) (view) Frank commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 10:05 PM. (go) (view) Dan Dare commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 09:02 PM. (go) (view) Leon Haller commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 08:52 PM. (go) (view) Helvena commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 08:48 PM. (go) (view) Leon Haller commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 08:42 PM. (go) (view) Graham_Lister commented in entry 'Review Call: Fighting for The Essence, by Dr Pierre Krebs' on 02/20/12, 08:38 PM. (go) (view) Guessedworker commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 08:36 PM. (go) (view) Graham_Lister commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 08:35 PM. (go) (view) dc commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 08:25 PM. (go) (view) Dan Dare commented in entry 'Kai Murros: Moscow Speech 2010' on 02/20/12, 08:23 PM. (go) (view) Recent Posts
General NewsHolocaust survivor's love story exposed as a fraud Patton was assassinated to silence his criticism of allied war leaders claims new book Nature on the ECHR's verdict on the surveillance society Ottawa fundraiser drops cystic fibrosis charity because disease only affects white males Scottish Government to fund school Auschwitz visits More Jews than ever sent to Washington Israeli Rahm Emanuel is Obama's chief of staff One of last remaining white-owned farms in Zimbabwe seized Spanish Queen speaks for her people on gay pride and gay marriage Interesting article on the Indian diaspora Blacks and Hispanics who suffer weight discrimination may perceive it as racial Horst Mahler goes on trial for Holocaust denial Roman invasion beach found in Kent Russian supreme court rules that Nicholas II and family must be recognised as victims Frattini's Blue Card progresses EJP reports assault on French Jews, mentions number but not race of attackers. Why might that be? Russian archaeologists find long-lost Jewish capital Sex industry "in every corner of London" Ron Paul's 'counter-convention' America's southern neighbour is succumbing to bloody anarchy Stonehenge was hidden by a 2-mile timber screen 100,000 young men missing from Australia Skull of mystery European woman who died in NZ a quarter of a century before James Cooke arrived Alpine melt reveals Neolithic life White victims don't always get a reliable physical picture of non-white perps Multiculturalism sank the Mary Rose Zut alors! Modesty back in fashion on the French Riviera 14th Century "newfangleness" stays in Britain Independent media activists fight back against Big Telecom's two-speed plan Young Muslims living in Britain turning to extremism Ratings of Bush, Cheney, Rice and Congress Sink to Worst Levels Ever Sweden's legislators push on with internet snooping bill British warship HMS Ontario found intact in Great Lake Tridentine Mass to return to England and Wales French to block porn, terror, hate web sites UCL prof claims working classes lack intelligence to be doctors White farming couple beaten and kicked off land in Zim Brigitte Bardot fined for inciting racial hatred Swiss don't feel threatened enough to vote for their own salvation Orthodox Jewish youths burn New Testaments Israel 'committing memorycide' The day finally arrives when Africa boasts more billionaires than the rest of the world together Nationalists vs. internationalists in Serb election Jewish Chronicle lists top 100 Jewish power wielders in UK Germany bans two incorrect groups Irish Viking trade centre unearthed Italian fascism on the rise again. Supposedly. "Waltzing Matilda" just a love song, not a socialist anthem Historian wants to resurrect the manly man Le Pen flogs his wheels to help indebted FN 30 girls contract TB in a Birmingham, UK Muslim school Zionists prove appeasement doesn't work Anecdotal evidence of impotence among teenage potheads Imported cheerleaders cause consternation in Indian cricket "Too moody" David Irving loses in court again - to his landlady Lupine ADL demand for curbs on free expression on-line US Senate blocks genetic discrimination Lynchings in Congo as penis theft panic hits capital Al-Zawahri says Shiite Iran spread the conspiracy theory that Israel was behind Sept. 11 attacks Poland tries to lure back UK emigrants A guy called Cohen is keen on white American guilt UKIP, Britain's anti-EU party, acquires a Member of Parliament Honda satnav warns when you drive in to high crime neighbourhood Calzaghe pays back Hopkins: "What's it like to get your arse kicked by a white guy?" Australia's new PM ingratiates himself with the intellectual and media elites Muslims want GMT replaced by Mecca time Britons fear race violence (BBC/Ipsos MORI poll) Zim's exiled white farmers working again - in Nigeria French government wants illegals out. Business and unions do not. TiVo tells the ad industry exactly how it's ignored, given the chance Darwin's first drafts of theory of evolution go online State security robots a decade away? Politicheskaya Pravil'nost reappears in not-Putin's Russia Bardot on trial for fifth time for inciting racial hatred against Muslim invaders Portrait of the disfunctional underclass No Eastern European crime wave according to police race and diversity head Capitol Hill to hear a whole lot more about Israel Italy returns Berlusconi with the support of Northern League Huaynaputina - the volcano that changed the world
(Entries from the original Majority Report home page have been archived below.) Richard Dawkins: Jews monopolize American policy (go man!) Sex trade of white women booming in Israel Jews have turned Bobruisk into a pigsty Man of the day (How a man with balls looks like) Christians in Jerusalem want Jews to stop spitting on them The SPLC admits it: Hal Turner an FBI agent (wise whites figured it out before) Lawmen under siege along Mexico border Science NewsTwo critical questions that remain unanswered: 1) How does one come to identify with a group? 2) What are the environmental triggers of oxytocin?
Milgram replicated: perfectly ordinary people can all-too-easily become torturers Same genes may govern intelligence and sperm quality Remains of Copernicus' identified Neural pathways spectacularly imaged by MRI Prehistoric cave paintings took up to 20,000 years to complete Tidy conservatives, messy liberals? First dig for 44 years dates Stonehenge at 2300 BC The face of Neanderthal woman revealed Brains and the taxi driver - or, maybe, why so many Jews used to drive London's black cabs Why we are evolved to be superstitious Study suggests AIDS-resistant gene less common where Romans ruled Women and property are the cause of male aggression Facial roundness in the male correlates with aggression. Whadd'ya know, I'm not a Neaderthal after all The Pill may change women's choice in men The Neanderthal murder mystery German villagers share DNA with cavemen US researcher discovers link between voting booth location and voting preference Sexual orientation nature, not nurture Gene evidence supports "Out of Africa" 200,000 years ago Conservatives Happier Than Liberals Women's voices more attractive during ovulation Human line split in Africa long before migrations north Japanese researchers find evidence that the need to belong is "biologically coded" Analytical technique shows how metabolism varies between populations James Watson's genome sequenced , and “It was so profound, how little we were actually able to say" Nationally representative American study finds no difference in testosterone levels between black and white men. The latest on racial differences in testosterone. Racial differences in eye anatomy Sex differences in sex drive, sociosexuality, and height across 53 nations Penis review: smaller in Asians but no clear differences among other races Self-pleasing behavior in Britain vs. China National IQs predict differences in scholastic achievement in 67 countries. 650,000 genetic markers and inferred population structure. Ethnocentrism increases during the first trimester of pregnancy. Important research on the nature of prejudice. Swedish men more masculine than Greenland Inuit. “Ethnic identity” predicts experimental pain sensitivity. Make that racial differences. Science CategoriesAll CategoriesThe WritersEach author's name links to a list of all articles posted by the writer; the hashes link to authors' homepages. LinksEndorsement not implied. Controlled Opposition Crime General
Immigration
Islam Jews
Nationalist Political Parties
Science Whites in Africa |
![Racial Kinship Coefficients of Nine Geographical Races X 10,000 (These show the kinship of random individuals from within a race in contrast to another race. Random co-ethnics have zero kinship when the ethnic group is taken in isolation. [Based on FST distances provided by Cavalli-Sforza et al., 1994, p. 80; rounded to nearest integers; standard errors omitted].)](egi_files/browse_003.gif)